
Техническое задание на проведение научно-исследовательской работы
Тема НИР: Исследование состояния нервно-мышечного аппарата и сенсомоторной системы спортсменов различных видов спорта.
Характер НИР: фундаментальное исследование
Руководитель НИР: Владимир Александрович Никишкин, заведующий кафедры физического воспитания и спорта МФТИ, кандидат физико-математических наук.
Вуз (организация), в котором проводится НИР: ГОУ ВПО «Московский физико-технический институт (государственный университет)»
Наименование структурного подразделения вуза (организации), в котором проводится НИР: Кафедра физического воспитания и спорта
Этапы НИР
| № Этапа | Наименование этапа | Сроки проведения | Научные и (или) научно-технические результаты (продукция) этапа |
|---|---|---|---|
| 1 | Разработка тестирования состояния нервно-мышечного аппарата и сенсомоторной системы человека. | 2009 г. | Описание состояния нервно-мышечного аппарата и сенсомоторной системы. |
| 2 | Разработка системы контроля и коррекции управления движениями в различных видах спорта. | 2010 г. | Описание системы контроля и коррекции управления движениями. |
Отчет по теме № 14.09 «Исследование состояния нервно-мышечного аппарата и сенсомоторной системы спортсменов различных видов спорта»
Введение
Соревновательные движения в спорте характеризуются целесообразностью, поэтому из огромной вариации возможных решений спортсмены находят наиболее выгодный. Выгода может быть выражена различными целевыми функциями, как, например, минимум времени, минимум затрат энергии, максимум силы и др. Поиск целевых функций (критериев) движения выполнялся в работах отечественных (П. К. Анохин, Н. А. Бернштейн, С. В. Голомазов, В. М. Зациорский, Р. С. Персон, Н. К. Рокотова, В. С. Фельдман и др.) и зарубежных ученых (P. M. Fitts, H. V. Komi, E. C. Poulton и др.). В результате оказалось, что любой разумный критерий дает совпадающие результаты между предсказанной электрической активностью мышц и реальной. Это означает, что решение пока не найдено, поэтому поиск механизмов управления двигательными действиями остается актуальной задачей. В связи с этим была определена цель работы — исследовать особенности работы нервно-мышечного аппарата и сенсорных систем при выполнении спортсменами определенных тестовых технических действий.
Объект исследования — спортсмены, выполняющие технические действия.
Основными методами являются анализ литературных источников, стабилография, электромиография, математическая статистика.
На первом этапе выполнен анализ литературных источников, определены основные достижения в области изучения биомеханики двигательных действий, особенности работы центральной и периферической нервной системы, педагогические и психологические особенности формирования двигательных навыков.
Биомеханика — как способ познания эффективности двигательных действий
Биомеханика движений человека необходима для понимания сущности двигательных действий, целесообразности построения движения. Она получила существенное ускорение в развитии в связи с научной деятельностью В. Брауне и О. Фишера (1897), Н. А. Бернштейна (1947, 1966).
Заслуга немецких ученых В. Брауне и О. Фишера заключается в том, что они впервые в биомеханике поставили задачу нахождения сил и моментов сил в центрах масс звеньев человека и практически решили ее. На трупном материале измерили массы и моменты инерции сегментов тела человека, с помощью метода хроноциклографии зарегистрировали кинематику движения, а затем оценили действующие силы при движении человека на основе Ньютоновской механики.
Н. А. Бернштейн повторил работу Брауне и Фишера, при этом он внес значительные усовершенствования в методику регистрации масс инерционных характеристик (МИХ) и кинематических характеристик. Далее он приступил к созданию физиологии двигательной активности, при этом последовательно разработал основы теоретической биомеханики, физиологии построения двигательного действия и теории технической подготовки спортсменов.
В очерке «О построении движений» Н. А. Бернштейн рассматривает двигательную систему позвоночных как модель (систему), состоящую из пассивной части (жесткий сочлененный скелет) и активной части (поперечно-полосатая мускулатура со всем ее оснащением). Затем дается описание свойств элементов (костей, связок, сухожилий, мышц), структурных образований (суставов, скелетно-мышечных моделей и др.).
Таким образом, Н. А. Бернштейн был одним из первых биомехаников, которые поставили в качестве краеугольного камня теоретической биомеханики строение опорно-двигательного аппарата, а свойства его использовались для объяснения причин определенной организации двигательного действия. Однако метода анализа двигательных действий Н. А. Бернштейн не создал. Изучение ходьбы и бега свелось только к эмпирическому анализу и представлению зарегистрированных данных, причем с явной ошибочной интерпретацией мелких «бугорков» на кривых «сила-время», поскольку артефакты принимались за закономерное явление — сенсорные коррекции.
Современное представление об анализе движений (техники) связано с понятием — биомеханизм (В. Н. Селуянов с соав., 1993, 1997, 2001).
Биомеханизм как основа теоретической биомеханики
Этапы анализа техники в теоретической биомеханике
Биомеханика движений человека граничит с физиологией двигательной активности Н. А. Бернштейна (1996), динамической морфологией и теоретической механикой. Для того, чтобы показать различие между биомеханикой и физиологией (нейрофизиологией) двигательной активности, приведем соображения Спинозы в интерпретации Э. В. Ильенкова (1974). «...Человеческая рука может совершать движение и по форме круга, и по форме квадрата, и по форме любой другой фигуры, обнаруживая тем самым, что структурно анатомически она заранее не предназначена к какому либо одному из названных действий «Человек — мыслящее тело — строит свое движение по форме любого другого тела». Следовательно, в мозгу человека могут формироваться любые программы двигательных действий, выполнение которых ограничивают пространственные, силовые и скоростные возможности опорно двигательного аппарата.
Программы движений, образы граничных поз являются идеальными представлениями. В природе нет идеального, однако человек тренирует органы своего тела на предметах окружающей среды, поэтому среда, его тело, способы управления закодированы в виде нервно-мозговых структур, т. е. вполне материально. Идеальное есть, следовательно, субъективное бытие предмета, или инобытие предмета в другом (мозг человека). На основе этих гносеологических положений можно утверждать, что человек представляет себя как часть пространства и мыслит перемещения своего тела в модели внешнего пространства. Он не знает анатомии и не может сознательно управлять определенными мышцами ради достижения поставленной цели.
В биомеханике можно не знать устройство мозга, достаточно иметь представление о ЦНС как о «черном ящике». Здесь проходит граница между физиологией и биомеханикой. Биомеханик — педагог должен овладеть методом программирования произвольных двигательных действий, обеспечивающих достижение заранее заданной цели. Такую же цель преследует тренер, когда пытается понять технические ошибки своего ученика и дать такие директивы, применение которых должно привести двигательные действия к желаемому виду.
На основании этих методологических положений можно определить объект и предмет исследования теоретической биомеханики движений человека:
Объект опорно двигательный аппарат человека (моделируется в виде скелета, мышц) и ЦНС.
Предмет закономерности построения моделей ОДА и программ управления ими в сознании человека, обеспечивающих выполнение произвольных и целенаправленных двигательных действий.
Среда — трехмерное пространство и материальные тела, с которыми взаимодействует объект исследования (человек).
Моделирование ОДА человека требует использования идеальных моделей теоретической механики. В теоретической механике применяются модели: двух или трехмерное пространство, время, материальная точка, абсолютно твердое тело (стержень), шарнир, кинематическая цепь, идеальная жидкость или газ и др. Все эти модели применяются в биомеханике, но для получения адекватной модели ОДА необходимо иметь модели мышцы и функциональной модели ЦНС различного уровня сложности (например, «черный ящик» всей ЦНС, который может иметь только один вход и выход). Следовательно, объект биомеханики пересекается с объектом теоретической механики, однако, не полностью. Если объединить вместе элементы ОДА и блок управления, то, согласно теории машин и механизмов, мы получим машину или механизм, с помощью которого можно производить какие-либо целевые преобразования.
В биомеханике эту конструкции логично обозвать биомеханизмом, поскольку включает элементы — мышцу и ЦНС, функционирующие по биологическим законам. Поэтому дадим следующее определение биомеханизму. Биомеханизм модель части или всего ОДА человека, функционирование которой, в соответствии с программой управления мышцами, обеспечивает достижение цели. В частности, повышает эффективность двигательного действия за счет преобразования одного вида энергии в другой, или способствует экономизации метаболической энергии, или способствует выходу максимума механической мощности без выхода за определенные физиологические критерии оптимальности функционирования организма.
Биомеханизм как целостная система состоит из совокупности элементов, входящих в ее состав. Каждый элемент обладает опреде-ленными свойствами, которые могут по-разному проявляется в движениях человека.
Можно выделить следующие элементы:
- Мышца как:
- преобразователь химической в механическую энергии;
- упругий элемент, способный накапливать и отдавать энергию;
- вязкий элемент, способный демпфировать внешние нагрузки;
- передатчик энергии (мощности) от других источников энергии.
- Кость как:
- рычаг для передачи силы или энергии;
- маятник для преобразования энергии;
- стержень для опоры и противодействия внешним нагрузкам.
- Сустав как:
- шарнир, соединяющий кости в кинематические цепи;
- шарнир, ограничивающий подвижность костей относительно друг друга.
- Блок управления мышцей (группой мышц-синергистов) как черный ящик, имеющий один вход и один выход. На вход поступает информация об интенсивности и начале выполнения упражнения, на выходе изменение относительной активности мышцы (мышц) во времени, другими словами, программа управления. Программа управления активностью мышцы (мышц) формируется человеком (или ученым для спортсмена) произвольно до начала движения.
Мышцы, кости, суставы, блок управления идеальные объекты теоретической биомеханики. Из этих конструктивных элементов человек создает из своего тела более или менее сложные системы биомеханизмы, с помощью которых достигается заранее поставленная цель движения.
Таким образом, развитие теоретической биомеханики приводит к стыковке ее с динамической морфологией и физиологией. Понятие биомеханизм становится объединяющим звеном. Конструкция биомеханизма — предмет анатомии, функционирование биомеханизма — предмет биомеханики и физиологии. Следовательно, теория в биомеханике есть описание биомеханизмов и способов достижения цели двигательного действия с помощью построения из тела человека и применения биомеханизмов во времени.
Теория управления двигательными дефствиями
Управление двигательными действиями осуществляется нервной системой, поэтому необходимо знать ее строение. На основе модели нервной системы можно разрабатывать инновационные педагогические технологии. Во второй части книги приводятся примеры таких технологий.Нейронная организация нервной системы
Нейрон является структурной единицей нервной системы. Он является клеткой подобно всем другим клеткам тела; однако определенные существенные отличия позволяют ему выполнять функции переработки информации и функции связи внутри мозга.
Как показано на рис. 1, нейрон состоит из трех частей: тела клетки, дендритов и аксона, каждая часть со своими, но взаимосвязанными функциями. Тело клетки управляет расходом энергии нейрона и регулирует множество других клеточных процессов.
Дендриты получают сигналы от других клеток через контакты, называемые синапсами. Отсюда сигналы проходят в тело клетки, где они суммируются с другими такими же сигналами. Если суммарный сигнал окажется большим порога, клетка возбуждается, дендриты и аксон передают импульсы на следующие клетки. Несмотря на очевидное упрощение, эта схема функционирования объясняет большинство известных процессов мозга.
Дендриты. Большинство входных сигналов от других нейронов попадают в клетку через дендриты, представляющие собой густо ветвящуюся структуру, исходящую от тела клетки. На дендритах располагаются шипиковые — синаптические соединения, которые получают сигналы от других аксонов.

Рис. 1. Нейрон
Синаптические контакты представляют собой узкое пространство, называемое синаптической щелью, отделяющее дендрит от передающего аксона. Специальные химические вещества, называемые нейротранс-миттерами, улавливаются специальными рецепторами на дендрите и внедряются в тело клетки. Тело клетки суммирует сигналы, полученные от дендритов и, если их результирующий сигнал выше порогового значения, вырабатывается импульс, проходящий по дендритам и аксону к другим нейронам.
Синаптическая связь, завершающая ветвь аксона, представляет собой маленькие утолщения, содержащие сферические структуры, называемые синаптическими пузырьками, каждый из которых содержит большое количество нейротрансмиттерных молекул — химических веществ возбуждающих мембрану клетки. Когда нервный импульс приходит по аксону, в синапсе некоторые из пузырьков высвобождают свое содержимое — медиатор в синаптическую щель, тем самым инициализируя процесс взаимодействия нейронов.
Аксон. Аксон может быть как коротким (0,1 мм), так и превышать длину 1 м. На конце аксон имеет множество ветвей, каждая из которых завершается синапсом, откуда сигнал передается на другие нейроны или клетки тела. Таким образом, всего один нейрон может генерировать импульс, который возбуждает или затормаживает сотни или тысячи других нейронов, каждый из которых, в свою очередь, через свои дендриты может воздействовать на сотни или тысячи других нейронов. Такая высокая степень связанности обеспечивает нейронной сети вычислительную мощность.
Нервный импульс
Информация поступает в нервные центры, перерабатывается там и затем передается эффекторам в виде нервных импульсов, пробегающих по нейронам и соединяющим их нервным путям.
Способ кодирования и передачи информации заключается в частоте следования нервных импульсов.
Независимо от того, какую информацию передают нервные импульсы, пробегающие по миллиардам нервных волокон, они ничем не отличаются друг от друга. Почему же в таком случае импульсы, идущие от уха, передают информацию о звуках, а импульсы от глаза — о форме или цвете предмета, а не о звуках или о чем-нибудь совсем ином? Да просто потому, что качественные различия между нервными сигналами определяются не самими этими сигналами, а тем местом, куда они приходят: в одной области мозга, в ней будет формироваться зрительный образ внешнего стимула, в другой — сигнал подвергнется расшифровке в виде, например, звуков. Теоретически достаточно было бы изменить ход нервных путей, например, часть зрительного нерва направить в зону мозга, ответственную за расшифровку звуковых сигналов, чтобы заставить организм «слышать глазами».
Нейронная сеть
Понятие нервной сети возникло на относительно ранней стадии исследований (приблизительно в середине ХIX в.), когда стало известно, что нервные клетки имеют длинные, сложно переплетающиеся отростки.
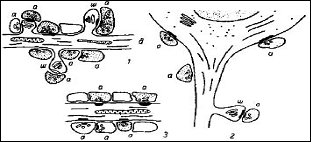
Рис. 2. Схема синаптических соединений (по Shepherd, 1979). 1-аксоношипиковый синапс; 2 — аксосоматический синапс; 3 — синаптические муфты и сериальные синапсы, а — аксон; д — дендрит; ш — шипик
Термин «нервная или нейронная сеть» встречали с подозрением до начала нашего века, когда нейрон начали считать основным клеточным элементом нервной ткани и ее анатомической единицей. Мы знаем теперь, что нервная ткань, хотя она построена из независимых клеточных элементов, соединенных специфическим образом и в специфических участках (синапсах), представляет собой скорее сеть сложно взаимосвязанных нейронов, чем их последовательные цепочки. Последовательные соединения, конечно, существуют, но не столько между отдельными нейронами, сколько между группами нейронов, соединенных рядом параллельных связей.
Считалось, что психические функции человека каким-то образом заложены в миллиардах корковых нейронов. Современный взгляд на мозг и психику совершенно иной. Кора оказалась вовсе не «автономной коллегией высших руководителей», многие области ее входят в состав сенсорных и двигательных систем, обеспечивая обработку информации на среднем уровне. Роль любой области коры зависит от внутренней организации ее синаптических сетей и от ее внешних связей с другими отделами — корковыми и подкорковыми. Организация синаптических сетей обуславливает те или иные функциональные свойства данного участка коры, а от его внешних связей зависит вклад этих свойств в функцию распределенной системы, в которую входит этот участок.
Значение коры для уникальных особенностей человеческой психики определяется организацией ее синаптических и шипиковых связей.
Микросети. Нижний уровень организации нейронных сетей представляют синапсы и шипики. Хотя синапс можно рассматривать лишь как простейший соединительный элемент сети, на самом деле — это сложная самостоятельная функциональная единица, в которой действуют регуляторные механизмы, зависимые от времени и интенсивности функционирования. Увеличение числа синапсов в коре, в процессе развития, в значительной степени связано с образованием шипиков.
Особенность коры головного мозга состоит в том, что шипики располагаются на дендритах в виде плотных цепочек, и это позволяет входным сигналам из разных источников взаимодействовать между собой. Можно представить себе, что вокруг каждого шипика создается микроучасток, в котором постсинаптический ответ изменяется в результате использования данной связи и влияет, прежде всего, на ответы соседних шипиков. Таким образом, каждый шипик действует как миниатюрный преобразователь сигналов, свойства которого зависят от предыстории его активности, от его метаболического аппарата, от входных сигналов и от взаимоотношений с соседними шипиками. Шипики имеют функциональное значение только в случае их соединения с телами или дендритами других нейронов.
Локальные сети. Синаптические сети коры, как и любого другого отдела ЦНС, построены из трех главных компонентов: входных нейронов, выходных нейронов и интернейронов.
Поскольку присущие человеку неповторимость определяет в основном неокортекс, то важно рассмотреть особенности, характерные для новой коры. Во-первых, эта кора расположена таким образом, что к ней могут поступать сигналы всех главных сенсорных модальностей. Они приходят сюда либо по прямым путям (от обонятельной коры), либо после переключения в стволе мозга и таламусе. Во-вторых, новая кора — это слоистая структура, образующая двойную складку; поэтому входные волокна идут в кору через глубинные слои, а выходные тоже проходят через эти слои. Благодаря такой особенности клетки всех слоев коры потенциально доступны для любых входных сигналов. В подобной структуре не доминирует какая-то определенная последовательность операций, подчиненная определенному входу или выходу. В-третьих, в коре имеется не один тип выходных нейронов, как во многих центрах, а несколько типов. По существу каждый слой коры служит источником выходных нейронов.
Локальные модули. Локальные сети обычно не распределены диффузно, а собраны в отдельные группы, или модули. В коре мозга имеются различные виды модулей. Модули не являются статичными, неизменными структурами.
Поля и доли коры. В каждой области коры многократно повторяются однотипные модули (локальные сети), благодаря которым данная область способна осуществлять специфические операции с участием определенных входных и выходных связей. При переходе к соседней области коры все эти три элемента — локальные сети, входы и выходы — несколько изменяются.
Каждое из таких полей есть участок, приспособленный для выполнения определенных функций в той распределенной системе, в состав которой он входит. Каждое поле отличается от соседних полей по функциональным свойствам и по связям в пределах такой системы.
Следующий уровень организации коры образуют ее доли. Кора больших полушарий у людей состоит из четырех главных долей: затылочной, теменной, лобной и височной. К затылочным долям приходят зрительные сигналы, к теменным — соматосенсорные, к височным — слуховые, а от лобных долей отходят многие двигательные пути.
В пределах каждой доли существуют поля, не связанные непосредственно с какой-либо сенсорной или моторной функцией, и их по традиции называют ассоциативными.
Имеется тесная связь между нейрохимическим составом участков коры и их структурной и функциональной дифференцировкой в пределах доли. Содержание и скорость оборота нейроактивных веществ в разных частях коры различны.
Было бы, конечно, удобно, если бы каждая доля коры отвечала за какую-то одну из высших психических функций, однако совершенно очевидно, что на самом деле это не так. В каждой доле содержатся центры определенных специфических функций; например, затылочная доля ответственна за зрительное восприятие. В то же время управление какими-либо навыками нельзя поместить в какой-то один центр. Даже при таком простом поведенческом акте, как захватывание предмета пальцами, координация движений руки и пальцев осуществляется с помощью зрения. Ясно, что для этого необходимы связи между зрительной корой, расположенной в затылочной доле, и центрами управления движениями в теменной и лобной долях.
Исследование поведения показало, например, что височные доли играют важнейшую роль в различении зрительных образов.
Согласно одной из концепций, каждую функцию обслуживает определенное сочетание корковых и подкорковых центров. Вклад каждого центра зависит от его функциональных особенностей, определяемых организацией его локальных сетей, а сами центры соединены между собой многочисленными длинными трактами, коллатералями и обратными связями.
Полушария. Передний мозг образован правым и левым полушариями, каждое из которых представляет собой совокупность всех долей соответствующей стороны. В настоящее время можно, пожалуй, утверждать, что левое полушарие обеспечивает определенные двигательные функции, тогда как правое в большей степени отвечает за глобальное представление о положении тела в пространстве, т.е. создает нечто вроде перцептивного "гештальта"- образа. То, что некоторые функции представлены только в одном полушарии, может означать, что это полушарие (обычно левое) подавляет активность другого. Иными словами, вследствие блокады недоминантного полушария доминантным через межполушарные волокна мозолистого тела недоминантное полушарие остается пассивным. Так, например, нервные сигналы, вызванные раздражением левой руки и приходящие в правое полушарие, автоматически передаются в доминантное левое полушарие. Лишь после того, как левое полушарие ознакомилось с этой информацией, в первое полушарие посылается команда, заставляющая левую руку выполнить нужное движение.
Модели управления движениями и формирования навыков
Скелетно-мышечная модель управляется мозгом, поэтому необходимо построить его модель. Работы Н.А. Бернштейна и П.К. Анохина положили начало построению моделей мозга, объясняющих процесс управления двигательными действиями. Идеи Н.А. Бернштейна и П.К. Анохина получили дальнейшее развитие в работах Н.К. Рокотовой. Модель мозга и периферии в ее варианте включает следующие блоки: 1) сенсоры и афференты, 2) нервную сеть сенсорных процессов, 3) блок моторных программ, 4) селектор программ, 5) адресный блок, 6) блок включения и развертки во времени, 7) блок двигательных нейронов, 8) мотонейроны спинного мозга (пул), 9) мышечные волокна. Каждый из блоков должен иметь свое место в ЦНС и строение, однако, до настоящего времени эта задача не решена. Исследования начала 21 века показывают, что при формировании двигательных программ активен мозг как целое. Поэтому представление о блоках лишь некая абстракция, решающая проблему в техническом исполнении, но не имеющая прямого отношения к работе мозга.
Моделирование центральной нервной системы
Предположения, высказанные Хеббом, ставшие теперь классическими, говорят о том, что: любые психические функции, будь то память, эмоции или мышление, должны быть обусловлены деятельностью нейронных ансамблей. Нервные клетки в таких ансамблях объединены в специфические сети. По мнению Хебба, при возбуждении нейрона его синаптические связи становятся более эффективными. Это может быть связано с кратковременным повышением возбудимости (кратковременная память) или же стойкими структурными изменениями в синапсах (долговременная память). Более корректно говорить о временном возникновении новых шипиковых связей.
Согласно Хеббу, следы памяти могут формироваться путем модификации синапсов (рис.4). На этой схеме показано, как два нейрона образуют синапсы на третьем. Сначала (I), когда активен нижний (темный) синапс, происходит возбуждение третьего нейрона, которое не может быть вызвано через слишком слабый верхний (светлый) синапс. Однако при одновременном воздействии обоих синапсов (II) третий нейрон тоже возбуждается, и в нем происходит ряд последовательных биохимических процессов, которые усиливают прежде слабый верхний синапс. В результате (III) верхний синапс приобретает способность сам по себе вызывать реакцию третьего нейрона.
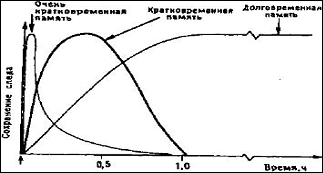
Рис. 3 Динамика следов памяти
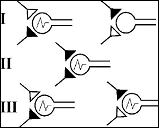
Рис. 4. Синапсы Хебба
Вот собственные слова Хебба: Предположим, что постоянная или повторяющаяся реверберация возбуждения (его «отзвук») вызывает в клетках долговременные изменения, которые усиливают стабильность такого процесса. Это предположение можно точно сформулировать следующим образом: «Если аксон клетки А расположен достаточно близко, чтобы возбудить клетку Б, и постоянно или многократно участвует в активации последней, то в одной или обеих клетках происходят процессы роста или обменные изменения, в результате которых усиливается активирующее действие клетки А на клетку Б». Согласно наиболее очевидному и, я полагаю, самому правдоподобному предположению о способе усиления возбуждающего действия одной клетки на другую, синаптические соединения разрастаются и увеличивают площадь контакта между пресинаптическим аксоном и постсинаптической клеткой. Разумеется, прямых доказательств правильности такого предположения нет. Однако по целому ряду соображений гипотеза о росте синапсов выглядит вполне приемлемой (Hebb D.O. The Organization of Behaviour, Wiley, 1949, рр.62-63).
Сейчас, когда известны закономерности образования шипиковых связей, особенности образования нейронных сетей стали более понятны. Среди нейробиологов наметилась тенденция в объединении трех наиболее обоснованных теорий памяти (Д. Адам). Согласно таким представлениям, электрические, синаптические и молекулярные процессы, вызываемые многократными внешними стимулами, зависят один от другого и протекают последовательно.
- Серия сенсорных импульсов, образующая частотный код, запускает реверберационную активность в самовозбуждающейся цепи. Реверберирующие бинарные сигналы, взятые в совокупности, сохраняют информацию в аналоговой форме. Эти следы памяти лабильны и быстро исчезают. Этот механизм памяти наименее вероятен.
- В случае очень интенсивных, часто повторяющихся, при сильной эмоциональной мотивации, стимулов ритмические изменения мембранного потенциала ускоряют внутриклеточный синтез белка. Это представление можно интерпретировать как образование множественных шипиковых связей. Закрепленные таким образом следы памяти весьма стабильны.
Биохимия памяти
Что нужно измерить, чтобы отличить нейрон, изменившийся после обучения?
Поверхность всех дендритов, отходящих от тела клетки, покрыта синапсами. Одни синапсы размещаются прямо на самих дендритах, другие на крошечных шипиках, отходящих от их поверхности.
Перестройка синаптических связей между нейронами в соответствии с гипотезой Хебба может быть связана с изменением длины дендритов, характера их ветвления или количества шипиков.
Расчеты биофизиков показали, что от синапсов, расположенных на шипиках, электрическая реакция распространяется эффективнее, чем от синапсов на самих дендритах, а также в отдельных шипиках, зависит от их формы. Характер межнейронных связей может изменяться не только при увеличении или уменьшении каждого отдельного синапса, есть веские основания полагать, что форма и характер ветвления дендритов имеют важное значение и могут изменяться под воздействием обучения или других форм приобретения опыта.
Процесс изменения моторных программ, а затем хранения этих изменений в памяти связан со структурными изменениями на клеточном уровне.
Во время работы любой моторной программы запускается каскад биохимических реакций. В результате происходит структурная модификация синаптических шипиковых связей, и в итоге это проявляется также в изменении электрических свойств клеток.
Биосинтез белков определяется информацией, заключенной в ДНК, т.е. в генах клеточного ядра. Гиперплазия органелл нейронов и содержимого дендри тов приводит к повышению функциональной активности нервных клеток, способствует более эффективному механизму функционирования шипиков, что ведет к облегчению образования шипиковых связей.
Значительная часть белкового синтеза связана с образованием новых шипиковых синапсов или модификацией старых, поэтому следует обращать внимание не на белки вообще, а на белки синаптических мембран. Многие из самых важных и характерных белков синаптических мембран относятся к классу гликопротеинов, которые можно описать как молекулы, состоящие из двух частей: длинной цепи аминокислот, погруженной в клеточную мембрану, и еще одной цепочки из молекул сахаров (например, глюкозы, фруктозы и галактозы), выступающей из мембран во внеклеточное пространство. Цепочки сахаров «липкие»: когда одна из них находит подходящую цепочку, выступающую над мембраной соседней клетки, они «узнают» друг друга и соединяются. Такими образом гликопротеины служат молекулами для узнавания. Скорость образования белков определяют, измеряя скорость включения аминокислот в белки.
Таковы современные представления о биохимических исследованиях памяти, корректность этих представлений еще надо проверять, пока существует простое объяснение явлений с помощью работы шипиков. В этом случае не требуется перестройки наследственного аппарата клетки.
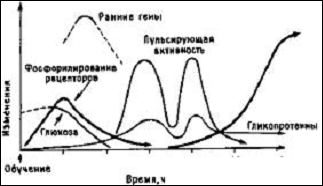
Рис. 5 Молекулярный каскад памяти. Кривые схематически показывают последовательность молекулярных изменений
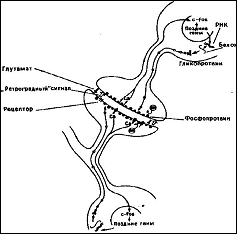
Рис. 6 Сигналы между синапсом и ядром. На рисунке изображен синапс шипик – шипикового соединения дендритов (без соблюдения масштаба)
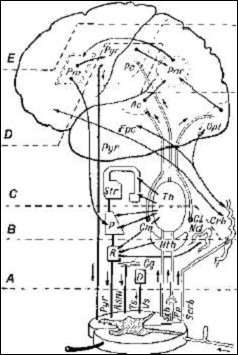
Рис. 7 Уровни построения движения (по Н. А. Бернштейну); символами обозначены ядра и проводящие пути мозга Н. А. Бернштейн (1966) полагал, что существует разные уровни организации двигательного действия. Экспериментально было показано, что такие уровни в ЦНС существуют, однако эти экспериментальные доказательства были связаны с оперативным вмешательством в нервную сиастему животного, с перерезкой иннервационных путей. В целом организме все нижележащие пути обратной связи вытормаживаются ради прямого управления из высших отделов ЦНС, так чтобы не было помех в достижении цели управления при произвольном программировании двигательной программы.
Процесс управления движением выполняется следующим образом. Например, регулирование размаха произвольного движения руки хорошо осуществляется как при наличии, так и в отсутствии зрительного контроля может быть свидетельством достаточности сигналов обратной связи только от опорно-двигательного аппарата. Когда размах движения назначается, его величина контролируется зрительной афферентацией. Выявлено, что каждому переносу руки на очередной финиш предшествует скачок глаз и фиксация взора на финальной позиции, промежуток времени составляет в среднем 206±21 мс. Смысл постоянного опережения взором руки заключается в сообщении «предварительной зрительной информации». После скачка глаз к очередному финишу имеется интервал времени (127±18 мс) прежде чем рука будет переведена со старта. В среднем фиксация взора на финишной позиции происходит на 839±13 мс раньше, чем рука переносится на финиш. Обнаружение постоянного опережения взором движения руки, при сохранении траектории серий движений позволило сделать вывод, что роль зрения не сводится к непосредственному контролю движений. Формирование в нервной системе указаний о направлении движения происходит как результат обработки зрительной информации о локализации объекта, относительно которого движение является целенаправленным.
Афферентные системы собственно двигательного аппарата передают информацию о внутренней стороне двигательной координации. В контроле движений они также реализуют две функции: задавание значения регулируемого параметра, и удержание параметра в заданных пределах.
Проведенные Н. А. Рокотовой опыты с введением тактильной коррекции в движения прицеливания показывают, что функция задавания свойственна и системам собственной чувствительности, формирующей так называемое «чувство позиции». Отличие проприоцептивной системы от зрительной системы автор видит в том, что для первой и «цель», и «рабочее пространство» связаны с функционированием двигательного аппарата.
При анализе механизмов афферентного контроля движений выделяют ряд последовательных этапов переработки информации — первичное описание параметров движения рецепторами, кодирование информации системами интернейронов спинного мозга (формирование вторичного описания), приём и переработка информации супраспинальными центрами, генерация афферентно обусловленных моторных ответов (как целостных координаций, так и сенсорных коррекций).
Теория формирования двигательных навыков
В основе обучения двигательным действиям лежит механизм запоминания, формирования следов в нейронных сетях, поэтому важно даже на эмпирическом уровне знать особенности функционирования этого механизма мозга человека.
Теоретические основы памяти
Большой интерес представляют данные о нейроанатомии, нейрогистологии и нейрофизиологии ЦНС (В. П. Бабминдра, Т. А. Брагина, 1982; Л. Н. Станкевич, В. П. Бабминдра, 1991). С морфологической точки зрения, нервные клетки подразделяются на звездчатые, пирамидные, веретеновидные и др. По количеству отростков их делят на униполярные, псевдоуниполярные, биполярные и мультиполярные нейроны. Дендритам принадлежит 80—90 % общего объема поверхности нейрона. На поверхности дендритов имеется множество шипиков, с помощью которых образуются дендро-дендрические электрические синапсы. Шипики представляют собой специфические выросты на дендритах и являются филогенически самыми молодыми образованиями в нервной системе. В онтогенезе они созревают значительно позже других структур; клетки и представляют собой ее наиболее пластический аппарат. Некоторые данные о структуре шипиков позволяют предположить, что они обладают как сенсорными, так и контрактильными механизмами. На поверхности выпуклой части шипика имеется синапс, как правило, возбуждающий. Подсчитано, что на пирамидном нейроне в коре человека содержится около 4400 шипиков, нейроны с шипиками занимают более 90 % клетки коры. Каждый аксон может образовывать мощный аппарат для объединения корковых модулей. Если образовалась шипиковая связь, при длительной потенциации шипики укорачиваются и становятся толще. Происходит притягивание дендритов друг к другу. Осуществляется это с помощью сократительных белков — стержневых актинсодержащих структур. Изменения формы шипиков могут составлять структурную основу памяти, поскольку при сближении дендритов возникает дополнительная возможность для возникновения новых шипиковых соединений. Так может преобразовываться кратковременная память в долговременную.
Эмпирические основы памяти
Память — форма психического отражения, заключающаяся в закреплении, сохранении и последующем воспроизведении прошлого опыта, делающая возможным его повторное использование в деятельности или возвращение в сферу сознания. Память связывает прошлое субъекта с его настоящим и будущим и является важнейшей познавательной функцией, лежащей в основе развития и обучения.
Память — основа психической деятельности. Без нее невозможно понять основы формирования поведения, мышления, сознания, подсознания, реализация двигательной деятельности. Поэтому для лучшего понимания процессов формирования двигательных навыков у спортсменов необходимо механизмы формирования памяти.
Амнезия — отсутствие памяти. Основные процессы памяти: заучивание, сохранение, воспроизведение, узнавание, забывание.
Выделяют следующие виды памяти:
- Непроизвольная память (информация запоминается сама собой без специального заучивания, а в ходе выполнения деятельности, в ходе работы над информацией). Сильно развита в детстве, у взрослых ослабевает.
- Произвольная память (информация запоминается целенаправленно с помощью специальных приемов). Эффективность произвольной памяти зависит от ряда причин:
- От целей запоминания (насколько прочно, долго человек хочет запомнить). Если цель — выучить, чтобы сдать экзамен, то вскоре после экзамена многое забудется, если цель — выучить надолго, для будущей профессиональной деятельности, то информация забывается меньше.
- От приемов заучивания. Приемы заучивания бывают нескольких видов:
- — механическое дословное многократное повторение — работает механическая память, тратится много сил, времени, а результаты низкие. Механическая память — это память, основанная на повторении материала без его осмысливания;
- — логический пересказ, который включает: логическое осмысление материала, систематизацию, выделение главных логических компонентов информации, пересказ своими словами — работает логическая память (смысловая) — вид памяти, основанный на установлении в запоминаемом материале смысловых связей. Эффективность логической памяти в 20 раз выше, лучше, чем у механической памяти;
- — образные приемы запоминания (перевод (информации в образы, графики, схемы, картинки) — работает образная память. Образная память бывает разных типов: зрительная, слуховая, моторно-двигательная, вкусовая, осязательная, обонятельная, эмоциональная; кроме того можно выделить следующие этапы логического запоминания:
- — мнемотехнические приемы запоминания (специальные приемы для облегчения запоминания).
- Выделяют также кратковременную память, долговременную память, оперативную память, промежуточную память.
Любая информация вначале попадает в кратковременную память, которая обеспечивает запоминание однократно предъявленной информации на короткое время (5-7 минут), после чего информация может забыться полностью либо перейти в долговременную память, но при условии 1-2 кратного повторения информации. Кратковременная память (КП) ограничена по объему, при однократном предъявлении в КП помещается в среднем 7±2. Объем кратковременной памяти у каждого человека индивидуален.
Долговременная память обеспечивает длительное сохранение информации. Она бывает двух типов: ДП с сознательным доступом (т. е. человек может по своей воле извлечь, вспомнить нужную информацию); ДП закрытая (человек в естественных условиях не имеет к ней доступа, а лишь при гипнозе, при раздражении участков мозга может получить к ней доступ и актуализировать во всех деталях образы, переживания, картины всей жизни человека).
Оперативная память — вид памяти, проявляющийся в ходе выполнения определенной деятельности, обслуживающий эту деятельность благодаря сохранению информации, поступающей как из КП, так и из ДП, необходимой для выполнения текущей деятельности.
Промежуточная память — обеспечивает сохранение информации в течение нескольких часов, накапливает информацию в течение дня, а время ночного сна отводится организмом для очищения промежуточной памяти и категоризации информации, накопленной за прошедший день, переводя ее в долговременную память. По окончании сна промежуточная память опять готова к приему новой информации. У человека, который спит менее трех часов в сутки, промежуточная память не успевает очищаться, в результате нарушается выполнение мыслительных, вычислительных операций, снижается внимание, кратковременная память, появляются ошибки в речи, в действиях.
Для долговременной памяти с сознательным доступом свойственна закономерность забывания: забывается все ненужное, второстепенное, а также определенный процент и нужной информации.
Для уменьшения забывания необходимо: 1) понимание, осмысление информации (механически выученная, но непонятная до конца информация забывается быстро и почти полностью); 2) повторение информации (первое повторение нужно через 40 минут после заучивания, т.к. через час в памяти остается только 50 % механически заученной информации). Необходимо чаще повторять в первые дни после заучивания, т. к. в эти дни максимальны потери от забывания, лучше так: в первый день — 2 — 3 повторения, в третий — седьмой день по 1 повторению, затем 1 повторение с интервалом в 7—10 дней. Следует помнить, что 30 повторений в течение месяца эффективнее, чем 100 повторений за день. Поэтому систематическая, без перегрузки учеба, заучивание маленькими порциями в течение семестра с периодическими повторениями через 10 дней намного эффективнее, чем концентрированное заучивание большого объема информации в сжатые сроки сессии, вызывающее умственную и психическую перегрузки и почти полное забывание информации через неделю после сессии.
Надо заметить, что этот механизм верен лишь для механической формы запоминания, для повышения эффективности запоминания используется множество приемов, в частности, построение моделей из приобретаемых знаний. В этом случае знания запоминаются не механически, а большая часть их воспроизводится с помощью имитационного моделирования.
Забывание в значительной степени зависит от характера деятельности, непосредственно предшествующей запоминанию и происходящей после нее. Отрицательное влияние предшествующей запоминанию деятельности получило название проактивного торможения. Отрицательное влияние следующей за запоминанием деятельности называют ретроактивное торможение, оно особенно ярко проявляется в тех случаях, когда вслед за заучиванием выполняется сходная с ним деятельность или эта деятельность требует значительных усилий.
Формы воспроизведения:
- — узнавание — проявление памяти, которое возникает при повторном восприятии объекта;
- — воспоминание, которое осуществляется при отсутствии восприятия объекта;
- — припоминание, представляющее собой наиболее активную форму воспроизведения, во многом зависящую от ясности поставленных задач, от степени логической упорядоченности запоминаемой и хранимой в ДП информации;
- — реминисценция — отсроченное воспроизведение ранее воспринятого, казавшегося забытым;
- — эйдетизм — зрительная память, долго сохраняющая яркий образ со всеми деталями восприятия.
На процессы памяти пытаются воздействовать также фармакологическими и физическими факторами.
Многие ученые считают, что поиски в области управления памятью должны быть направлены на создание биологически активных соединений, избирательно влияющих на процессы обучения (скажем, кофеин, биогенные амины), кратковременную или долговременную память (вещества, тормозящие синтез ДНК и РНК, влияющие на обмен белков и др.), создание и формирование энграмм — веществ, влияющих на изменение белков клетки (от протоплазмы до сомы).
Сейчас изучение фармакологических средств, влияющих на память, идет стремительно. Установлено, что давным-давно известные гормоны гипофиза могут служить стимуляторами памяти. «Короткие» цепочки из аминокислот — пептиды, особенно вазопрессин, кортикотропин значительно улучшают кратковременную и долговременную память.
Способность мозга определять время совершения события в основном зависит от памяти.
Память, очевидно, не связана с какой-то одной специфической областью мозга; она зависит от многочисленных зон, играющих важную роль. В особенности это касается некоторых областей височной коры и в еще большой мере — гиппокампа.
Планирование действий, которое, собственно, и составляет суть мышления, происходит в префронтальной коре (т. е. в передних участках лобных долей) в результате объединения и переработки ею информации, получаемой и расшифровываемой в других зонах коры.
Теория технически подготовки связана с исследованиями изменений или стабильностью выполнения так называемых «моторных программ». Процессы обучения и памяти, связанны с хранением, извлечением и изменением моторных программ.
Внутренний и окружающий мир спортсмена описывается с использованием представлений о схеме тела, чувстве движения, внешней ориентации и текущем состоянии.
Схема тела
Для реализации моторных программ мозг должен уметь формировать внутреннее представление об актуальном окружении (модель мира), а также иметь представление о собственном теле, его структурной организации, его сенсорных и моторных возможностях и т.п. (модель самого себя). В пользу предположения о том, что в процессах регуляции позы и движений участвует внутренняя модель тела, свидетельствуют накопленные в клинической и психологической литературе данные о так называемой «схеме» тела.
Например, о положении и движениях головы, конечностях и всего тела мы обычно узнаем без помощи зрения. Даже при закрытых глазах мы знаем, вытянута или согнута у нас рука или нога, повернута голова вперед или вбок, стоим ли мы прямо. Эту информацию о положении и движениях тела и его различных частей доставляет в мозг сенсорная система. Рецепторы этой сенсорной системы находятся в мышцах и суставных сумках, а также в вестибулярном аппарате внутреннего уха. Эти рецепторы получили название проприоцепторов; это значит, что они информируют нас о состоянии нашего собственного тела.
Не все элементы статической и кинетической информации (т. е. информации о положении и движениях тела) достигают большого мозга. Генерируемые в проприоцепторах импульсы могут активировать врожденные автоматизмы, приводящие к рефлекторной деятельности, которая регулируется не корой, а продолговатым или средним мозгом; иногда эти импульсы даже прямо проводятся к передним рогам спинного мозга. Некоторые движения, связанные с прямостоянием, совершаются автоматически. Реакции, связанные с тонким регулированием положения тела и головы, доходят до сознания с задержкой или же не доходят вовсе. Таким образом, некоторые процессы, происходящие в проприоцептивном анализаторе, остаются неосознанными. Та часть информации, которая осознается человеком, проходит посредством проприоцептивных импульсов по сложному пути, самовозбуждающемуся кольцевому пути, который включает сенсорные и моторные зоны коры, определенные группы нейронов в стволе мозга (анатомы назвали их базальными ганглиями), нейроны таламуса и мозжечковые структуры. Эту реверберационную систему, которая связана с управлением схемой тела, неврологи называют экстрапирамидной системой. Влияние мышечного утомления, какие-либо изменения в опорно-двигательном аппарате (например, изменение весоростовых показателей) будут оказывать прямое влияние на работу данной системы. Надо сказать, что уровни организации движения, мешающие произвольному выполнению задуманного двигательного действия, со временем вытормаживаются.
Чувство движения
Чувство движения — это способность представлять будущее движение или воспроизводить его в идеальной форме Человек может строить идеальные программы движения и затем их реализовывать, сначала с ошибками. Далее человек управляет, с большей или меньшей точностью, целенаправленными движениями всего тела или его отдельных частей.
Как пример, можно привести набор биомеханизмов, участвующих в осуществлении прыжка вверх с места: биомеханизм разгибания ног и туловища, биомеханизм свободного падения тела при незакрепленных суставах, биомеханизм маховых звеньев (В. Селуянов, Ан. Шалманов, 1995). Каждый из этих механизмов можно реализовать с разной и управляемой степенью интенсивности.
Цель движения решается при последовательном или одновременном действии конкретных биомеханизмов. В большинстве случаев имеют место, так называемые, препрограммируемые произвольные движения. Техника сначала разучивается, каждый раз воспроизводится в идеальном виде, а затем она используется в автоматическом режиме. Разучивание выполняется в форме последовательного планирования некоторых изменений сначала в идеальном виде, без физической активности, а затем в рельном — физическом, как правило, решение превращается в перепрограммируемое действие. Если изменения приводят к приближению формы движения к модели, то изменения в программе движения стараются закрепить (запомнить).
Внешняя ориентация
Ориентация во внешнем мире, привязка к месту и времени. Для выполнения даже такого простого двигательного акта, как захватывание предмета, необходима координация между зрительным и тактильным восприятием пространства. Для этого нужно, чтобы существовал участок (или участки), где интегрировались бы эти две модальности. По-видимому, важным участком, где интегрируются зрительные и тактильные восприятия, служит задняя область теменной коры.
Высказано предположение, что в гиппокампе содержится «когнитивная карта», в которой как бы отмечены места различных прошлых событий. Можно добавить, что в гиппокампе хранится комбинированная информация о месте и времени прошлых активных действий и о сопутствующих им эмоциях.
Как ориентация, так и пространственная память, связаны с деятельностью распределенных систем, включающих множество корковых (а также подкорковых) областей.
Текущая ситуация
Изменение обстановки, реагирование на возникающие помехи. Возможность принятия решения (выбор из набора имеющихся двигательных программ) или изменение имеющейся программы. Соответственно, существует проблема распознавания ситуации, которая также решается в процессе обучения распознаванию ситуаций.
Системная организация оказывается связанной с выбором предпочитаемых исполнительных механизмов, обеспечивающих возможность разных стратегий поведения при наличии одной и той же цели или в различных ситуациях. Из этого следует, что при наличии какой-либо уже встречающейся ранее цели или постоянной цели возможна активация не единственной программы, а выбор одной из нескольких имеющихся, но неактивированных программ.
Простейшая модель центральной нервной системы
Можно предположить, что в коре головного мозга имеется множество структурно оформленных блоков нейронов, отвечающих за выполнение определенных физиологических функций. Например, в двигательной зоне располагаются нейроны, управляющие активностью скелетных мышц. Активность такого нейрона описывается двумя переменными: А — активностью (число импульсов в секунду), и П — продолжительностью активности. Поскольку активация мышц, как правило, идет в импульсном режиме, то достаточно знать только эти два параметра.
Распределение активности нейронов во времени вызывает определенное рекрутирование ДЕ, с определенной активностью мышечных волокон. Внешне это выглядит как сохранение требуемой позы и реализация внешней формы движения определенными частями тела.
У человека имеются рецепторы, поэтому сигналы от них входят в сенсорный блок и фиксируются в виде шипиковых соединений — образуется в памяти след. Этот след человек способен воспроизводить уже без активности мышц, создавать ощущение выполненного двигательного действия. Также он может воспроизводить программу управления двигательными нейронами, но без их активации. Для этого в мозгу должен быть специальный блок нейронов куда посылаются и сравниваются как ощущения от выполненного действия, так и программа управления двигательными нейронами. Сопоставление этих следов и неоднократное проигрывание позволяет принять решение либо об адекватности их, либо формируются целевые установки по коррекции двигательной программы. Практическая реализация новой двигательной программы создает новые ощущения, новый процесс запоминания и сравнения с критериями эффективности разучиваемого движения. Таким образом, повторное выполнение разучиваемого двигательного действия должно создавать в блоке памяти программ множество структурных (шипиковых соединений) следов, которые будут относится ко всем видам попыток реализации, а именно, к очень плохим, похожим и правильным. При повторном исполнении движения шипиковые связи закрепляются и переходят из кратковременной к долговременной форме памяти. При частом повторении будут закрепляться все повторно выполняемые двигательные программы как правильные, так и неправильные.
Точность выполнения перепрограммируемого двигательного действия будет зависеть от исходной информации, выбора цели, наличия группы адекватных двигательных программ, корректности выбора нужной программы. При реализации двигательной программы результат будет зависеть еще и от состояния исполнительного аппарата, например, утомление может приводить к неадекватной реакции мышечных волокон на предъявляемые стимулы.
Педагогические основы формирования двигательных действий
Процесс технической подготовки проходит наиболее эффективно в случае формирования у спортсмена ориентировочной основы двигательного действия. В психологии благодаря работам Л. С. Выготского (1960), А. Н. Леонтьева (1977) стало возможным изучение процесса обучения как деятельности. Структурными компонентами деятельности выступают действия. Действием называют процесс, подчиненный решению конкретной задачи, направленный на достижение цели, определяемой характером мотивации. С позиций теории деятельности П.Я Гальпериным и его сотрудниками (1985) была разработана теория поэтапного управления процессом усвоения знаний. В этой теории образ действия и образ среды, в которой это действие происходит, объединяются в единую систему, на основе которой происходит управление действием, названным «ориентировочной основой действия» (ООД). ООД не тождественна той картине, которую видит внешний наблюдатель. ООД — это образ (модель), существующий только в сознании или в подсознании (субъекта), как в виде теоретического знания о последовательности действий, так и в виде реальной программы (пакета программ) движения. Контроль качества программ может осуществляться как путем проверки теоретических знаний по биомеханике данной техники, так и в результате анализа двигательной деятельности.
Успешность выполнения действия зависит от степени полноты ООД (полная — неполная), меры обобщенности (обобщенная — конкретная) и способа ее получения (самостоятельная — «в готовом виде»), как пишут П. Я. Гальперин (1985) и Н. Ф. Талызина (1984).
Сочетание каждого из тех или иных показателей по каждому из трех критериев по данным Н. Ф. Талызиной определяет тип ООД. Например, ООД может быть неполная, конкретная, получена самостоятельно (1 тип); ООД полная, конкретная, данная «в готовом виде» (2 тип); ООД полная, обобщенная, данная «в готовом виде» (3 тип). Всего возможны 8 типов. Соответственно трем главным типам ООД выделяют три типа учения.
Первый тип учения имеет место при уяснении схемы ООД первого типа. В этом случае уяснение и дальнейшая обработка проходят с ошибками, с недостаточным пониманием содержания усваиваемого материала и существенных признаков, их отличием от несущественных признаков в полном объеме, и всегда применительно к конкретному материалу. Если в относительно простых случаях занимающийся «усматривает» правильное решение задачи, то «сознательность остается невоспитанной, и не только ребенок, но и взрослый, искусно выполняющий действие, как утверждает П. Я. Гальперин, большей частью ничего не может о нем рассказать».
Второй тип учения имеет место при уяснении схемы ООД второго типа и характеризуется более уверенным и полным содержанием материала, с четким различением существенных и несущественных признаков понятий и состава действий, также применительно к ограниченной конкретной области.
Третий тип учения имеет место при уяснении схемы ООД третьего типа. Для данного типа учения характерно эффективное протекание процесса понимания содержания учебного материала, с уяснением существенных и несущественных признаков изучаемых явлений и условий действия с ними в широкой области, и самостоятельным переносом знаний о действиях на все конкретные случаи в пределах данной области.
В третьем типе учения различают два начала: метод исследования объектов и метод вовлечения в это исследование. Метод исследования объектов нацелен на выяснение основной структуры изучаемых объектов — основных единиц данной области и способов их сочетания в более крупные, конкретные образования. Метод вовлечения в исследование направлен на возбуждение познавательного интереса у занимающихся. Оба метода глубоко связаны. Результатами экспериментальных исследований доказано, что именно установка на изучение объекта помещает основную цель деятельности в самый процесс познания, раскрытия предмета, возбуждает познавательный интерес к нему и побуждает к использованию тех возможностей, которые открывает метод исследования.
Каждое действие содержит 3 части: ориентировочную, исполнительную и контрольно-корректировочную (М. М. Боген, 1985, 1987). В результате осуществления первой части формируется ООД, вторая часть реализует ту программу, которая формируется на основе ООД, это — попытка решить двигательную задачу; третья часть анализирует результат действия — контролирует качество первых двух, оценивая их по эффективности продвижения к цели. Если исполнительная часть не соответствует требованиям ООД, то в нее вносятся соответствующие коррективы. Если отклонений от ООД нет, но продвижение к цели неудовлетворительно, то коррективы вносятся в ООД. Третья часть действия подает сигнал о прекращении действия по достижению намеченного результата.
Все три части существуют в единстве, что позволяет рассматривать действие как систему управления, в которой управляющий орган — ООД, управляемый орган — исполнительная часть, следящая система — контрольно-корректировочная часть.
На практике имеет место обучение методом «проб и ошибок», при котором уяснение и дальнейшая обработка проходят с ошибками, с недостаточным пониманием содержания усваиваемого материала. Если в относительно простых случаях занимающийся «усматривает» правильное решение задачи, то «сознательность остается невоспитанной». Что касается методов тренировки точности двигательных действий, то анализ литературы показал, что существует вполне определенная точка зрения, согласно которой разработка методов ведется на основе физиологических положений И. П. Павлова о дифференцировочной деятельности коры головного мозга, а именно:
- многократное повторение одного и того же раздражителя еще не приводит к его специализации, дифференцировке от других, сходных раздражителей;
- дифференцирование осуществляется при чередовании двух близких или сначала отдаленных, а затем все более сближающихся по характеру раздражителей.
Благодаря такому пониманию выделяют следующие методические приемы:
- прием «смежных» заданий — в котором выработка дифференцировок осуществляется при чередовании двух близких по характеру раздражителей;
- прием «контрастных» заданий — чередуются отдаленные раздражители;
- прием «сближаемых» заданий — предполагает постепенный переход от «контрастных» к «смежным» заданиям.
При методе смежных заданий выработка дифференцировок осуществляется при чередовании двух близких по характеру раздражителей.
В основу метода сближаемых заданий положено использование так называемого дифференцировочного торможения. Те раздражители, которые поначалу воспринимаются как одинаковые, постепенно начинают различаться. Предусматривается выполнение одних и тех же заданий, сначала резко отличающихся друг от друга по величине раздражителя (стимула), с постепенным сближением величины раздражителя в последующих попытках.
Контрастный метод в некоторой степени схож с методом сближаемых заданий. При тренировке контрастным методом предусматриваются задания, отличающиеся друг от друга, но со сходной структурой движений выполнения приема. По мнению С. В. Голомазова, наибольший эффект достигается в том случае, когда контрастные задания составляют не более 25 % от общего объема.
Описанные выше методы тренировки точности используются в виде различных методических приемов. Наиболее распространенными приемами для тренировки целевой точности являются следующие: варьирование веса снаряда, включение или выключение зрения, варьирование размеров цели, дополнительные ориентиры.
Предполагается, что варьирование веса снаряда как некоего стимула должно изменить двигательную афферентацию. В свою очередь, изменение величины раздражителя должно отразиться на точности движений.
Применение утяжелённых мячей не дает положительных результатов ни с точки зрения совершенствования техники, ни с точки зрения точности, а в ряде случаев приводит к отрицательному результату.
Выключение зрения как средство тренировки точности баскетбольных бросков способствует улучшению точности, снижению ее погрешностей как в дальности так и в направлении бросков. При тренировке баскетболистов этот прием позволил достичь более значительного прироста точности, чем обычные броски.
Заключение
Анализ литературных источников показал, что проблема изучения информационных процессов в теле спортсменов остается далекой от своего разрешения. Существует разные стороны изучения техники движений:
- — исследование внешней формы движений с помощью биомеханических методов,
- — исследование процессов в нервной системе с помощью электромиографии,
- — исследование процессов формирования двигательных навыков с помощью нейронных сетей.
Для исследования основных закономерностей реализации двигательных навыков и формирования их необходимо использовать комплексные методики.
Стабилография, электромиография, кардиография — все эти методы объединены в СТАБИЛАНЕ. В связи с этим на втором этапе выполнения работ по теме 14.09 будут решаться следующие задачи:
- — исследовать особенности функционирования нервно-мышечного и сенсорного аппарата при выполнении тестовых заданий на стабилографе,
- — исследовать двигательные программы сохранения равновесия у спортсменов различных видов спорта.
Список использованной литературы
- П. К. Анохин. Опережающее отражение действительности // Вопросы философии. — 1962. — № 7. — С. 30 — 36.
- П. К. Анохин. Биология и нейрофизиология условного рефлекса. — М.: Медицина, 1968. — 117 с.
- И. П. Башлыков, М. Х. Казиев. Оценка точности двигательных действий // Лекция для студентов, аспирантов и слушателей факультета повышения квалификации ГЦОЛИФК. — М., 1989. — С. 3 — 4.
- Н. А. Бернштейн. О построении движений. — М.: Медгиз. — 1947. — 380 с.
- Н. А. Бернштейн. Очерки по физиологии движений и физиологии активности. М.: Медицина. — 1966. — 349 с.
- Биомеханизмы как основа развития биомеханики движений человека (спорта) / В. Н. Селуянов, Ан. А. Шалманов, Аиед Берхаием, К. А. Анненков, А. В. Григоренко // Теория и практика физической культуры. — 1995. № 7. — С. 6 — 10.
- М. М. Боген. Методологические основы теории обучения двигательным действиям. Учебное пособие для слушателей факультетов усовершенствования и аспирантов / ГЦОЛИФК, М., 1985. — 38 с.
- М. М. Боген. Современные теоретико-методические основы обучения двигательным действиям: Автореф. дисс. ... д-ра пед. наук. — М., 1989. — 37 с.
- В. Н. Селуянов. Теория и практика дидакттики развивающего обучения в подготовке специалистов по физическому воспитанию: Труды сотрудников проблемной научно-исследовательской лаборатории РГАФК. / Научный ркуководитель В. Н. Селуянов. — М.: Физкультура, образование и наука, 1996. — 106 с.
- Н. Винер. Кибернетика или управления и связь в животном и машине. — М.: Сов. радио, 1958. — 215 с.
- Л. С. Выгодский. Развитие высших психических функций. М., АПН, 1960. — 55 с.
- П. Я. Гальперин. Методы обучения и умственное развитие ребенка. — М.: изд-во Моск. ун-та, 1985. — 45 с.
- С. Г. Геллерштейн. Действия, основанные на предвосхищении и возможность их моделирования в эксперименте // Материалы по проблемам инженерной психологии. — М., 1966. — Вып. 6. — С. 142 — 154.
- С. Г. Геллерштейн. Чувство времени и скорость двигательной реакции. — М.: Медгиз, 1958. — 148 с.
- Г. И. Гинсбург. Основные понятия, критерии оценки и классификация точностных движений / Г. И. Гинсбург, А. В. Ивойлов, В. А. Левчук, В. М. Замота // Точностные движения в спортивных играх. — Волгоград, 1986. — С. 9 — 12.
- Ю. Б. Гиппенрейтер. Движение человеческого глаза / МГУ. — М., 1978. — 185 с.
- С. В. Голомазов. Исследование механизмов управления точностью движений и экспериментальное обоснование методики ее повышения: Автореф. дис. ... канд. пед. наук. — М., 1973. — 30 с.
- С. В. Голомазов. Состояние исполнительного аппарата как фактор, определяющий надёжность целевого препрограммируемого двигательного действия / С. В. Голомазов, М. М. Кадри, В. Н. Селуянов, М. Шейх. // Теория и практика физической культуры. — 1994. — № 4. — С. 27 — 30.
- С. В. Голомазов, Зациорский В. М. Точность двигательных действий: Учеб. пособие для студентов ин-тов физ. культуры / ГЦОЛИФК. — М., 1979. — 26 с.
- С. В. Голомазов. Исследование механизмов управления точностью движений и экспериментальное обоснование методики ее повышения (на примере баскетбольных бросков): Автореф. дисс. ... канд. пед. наук. — М., 1973. — 24 с.
- С. В. Голомазов. Теоретические основы и методика совершенствования целевой точности двигательных действий: автореф. дисс. ... д-ра пед. наук. — М., РГАФК, 1997. — 42 с.
- С. В. Голомазов, Б. И. Чирва. Футбол. Тренировка точности юных спортсменов. — М., 1984. — 81 с.
- В. В. Давыдов. Проблемы развивающего обучения. Опыт теоретического и экспериментального психологического исследования. — М.: Педагогика, 1986. — 240 с.
- В. М. Дьячков. Экспериментальное обоснование и разработка системы тренировки в скоростно-силовых видах спорта: Автореф. дис. ... д-ра пед. наук. — М., 1963. — 26 с.
- В. М. Зациорский, С. В. Голомазов, М. Х. Казиев. Успешность приема мячей в зависимости от скорости, направления и длительности полета // Теория и практика физ. культуры. — 1984. — № 8. — С. 25 — 27.
- В. М. Зациорский, П. Р. Казаков, Г. А. Смирнов. Факторы, влияющие на точность ударов футболистов // Теория и практика физ. культуры. — 1975. — № 5. — С. 15 — 20.
- В. М. Зациорский, А. С. Аруин, В. Н. Селуянов. Биомеханика двигательного аппарата человека. М.: Физкультура и спорт, 1981. 143 с.
- Э. В. Ильенков. Диалектическая логика. — М.: Политиздат, 1984. — 320 с.
- Е. П. Ильин. Асимметрия точности движения рук в пространстве. — В кн.: Материалы IX Всес. науч. конф. по физиол., морфол. И биомеханике мышечной деятельности. М., 1966, т. 2, с. 12 — 13.
- Е. П. Ильин. Влияние многолетней односторонней тренировки на степень выраженности функциональной асимметрии. — Теория и практика физической культуры, 1961, т. 24, № 3, С. 200 — 203.
- Е. П. Ильин. Некоторые особенности проявления мышечного чувства, связанные с функциональной асимметрией. — Теория и практика физической культуры, 1966, № 5, с. 60 — 62.
- Е. П. Ильин. О симметрии и асимметрии в деятельности двигательного анализатора человека.: Автореф. дис. ... канд. биол. наук. — Л., 1962. — 21 с.
- М. Х. Казиев. Двигательные действия, связанные с реакцией на движущийся объект (летящий мяч), и некоторые пути повышения их эффективности: Автореф. дис. ... канд. пед. наук. — М., 1989. — 22 с.
- В. С. Келлер. Деятельность спортсменов в вариативных конфликтных ситуациях. — Киев: Здоровь’я, 1977. — 184 с.
- В. С. Келлер. Исследование деятельности спортсменов в вариативных конфликтных ситуациях: Автореф. дис. ... д-ра пед. наук. — Львов, 1974. — 48 с.
- В. Б. Коренберг. Качественный кинезиологический анализ как педагогическое средство в спорте: Автореф. дисс. ... д-ра пед. наук. — Малаховка, 1995. — 57 с.
- А. Н. Леонтьев. Деятельность. Сознание. Личность. / А. Н. Леонтьев. — 2-ое изд. — М.: Политиздат, 1977. — 304 с.
- Б. Ф. Ломов, Е. Н. Сурков. Антиципация в структуре деятельности. — М.: Наука, 1980. — 277 с.
- Г. П. Лукирская. Биомеханическое исследование структуры сложно-координационного ударного движения в процессе его формирования: Автореф. дисс. ... канд. биол. наук. — Л., 1968. — 23 с.
- В. П. Лукъяненко. Точность движений, проблемные аспекты теории и их прикладное значение // Теория и практика физической культуры. — 1991. — № 4. — С. 2 — 10.
- Л. Е. Любомирский. Управление движениями у детей и подростков. — М.: Педагогика, 1974. — 232 с.
- Л. П. Матвеев. Основы спортивной тренировки: Учеб. пособие для ин-тов физ. культуры. — М.: Физкультура и спорт, 1977. — 280 с.
- Л. П. Матвеев. Теория и методика спорта: Учеб. материалы / Л. П. Матвеев: ГЦОЛИФК. — М., 1992. ч. 1. — 224 с.
- Л. П. Матвеев. Общая теория спорта: для завершающих уровней высш. физ. образования. — М.: (4-ый фил. Воениздата), 1997. — 304 с.
- Моторные задачи и исполнительная деятельность. Исследование координированных движений руки / Н. К. Рокотова, Е. К. Бережная, И. Д. Богина, И. М. Горбунова, Е. С. Роговенко. — Л.: Наука, 1971. — 180 с.
- Р. С. Персон. Мышцы антагонисты в движениях человека. М.: Наука, 1965. — 115 с.
- Р. С. Персон. Электромиография в исследованиях человека. — М.: Наука. 1969. — 231 с.
- В. Н. Платонов. Адаптация в спорте. Киев: Здоров’я, 1988. 216 с.
- В. Н. Платонов. Теория и методика спортивной тренировки. Киев: Вища школа, Голов. изд-во, 1984. 352 с.
- А. А. Поцелуев. Роль спортивных игр в улучшении функции левой руки у школьников.: Автореф. дисс. ... канд. пед. наук. — Л., 1951. — 21 с.
- А. В. Родионов. Психодиагностика спортивных способностей. — М.: Физкультура и спорт, 1973. — 216 с.
- А. В. Родионов. Психофизическая тренировка. — М.: [ТОО Дар], 1995. — 64 с.
- А. В. Родионов. Спортсмен прогнозирует решение. — М.: Физкультура и спорт, 1971. — 71 с.
- Н. К. Рокотова. Моторные задачи и исполнительная деятельность. Исследование координтрованных движений руки / Н. К. Рокотова, Е. К. Бережная, И. Д. Богина, И. М. Горбунова, Е. С. Роговенко — Л.: Наука. 1971. — 180 с.
- P. H. Рудаков, Ю. И. Няшин, А. Р. Подгаец, А. Ф. Лисовский, С. А. Михеева. Влияние аэродинамических сил на движение спортсменов и спортивных снарядов // Российский журнал биомеханики / Т. 5, № 2. — С. 83 — 94.
- В. Н. Селуянов. Направления развития теории технической подготовки спортсменов // Проблемы теории технической подготовки спортсменов. — М.: ГЦОЛИФК, 1993. — 45 с.
- В. Н. Селуянов, Аиед Берхаим. Биомеханизм как основа развития теоретической биомеханики двигательной деятельности человека. Учеб. пособ. для студентов и слушателей РГАФК. — М., 1997. — 82 с.
- Е. Г. Сироткин, В. Н. Селуянов. Совершенствование техники исполнения прыжка с вышки из задней стойки с вращением вперёд // Проблемы теории технической подготовки спортсменов. — М., ГЦОЛИФК, 1993. — С. 35 — 36.
- Г. А. Смирнов. Исследование факторов, влияющих на меткость ударов футболистов, и некоторые пути ее воспитания: Автореф. дис. ... канд. пед. наук. — М., 1975. — 23 с.
- В. В. Соломонко. Тренировка вратаря в футболе. — Киев: Здоров’я, 1986. — 128 с.
- Спортивная физиология: Учебник для ин тов физ. культуры / Под ред. Я. М. Коца. М.: Физкультура и спорт, 1986. 240 с.
- Структурные основы адаптации и компенсации нарушенных функций.:Руководство / Л. И. Аруин и др. Под ред. Д. С. Саркисова. М.: Медицина,1987. 448 с.
- Е. Н. Сурков. Антиципация в спорте. — М.: Физкультура и спорт, 1982. — 144 с.
- Е. Н. Сурков, Ю. С. Якобсон. Пути обучения футбольных вратарей упреждающим действиям // Научные основы физического воспитания и спорта. — Л., 1976. — С. 104 — 105.
- Н. Ф. Талызина. Психологические основы программированного обучения. — М., 1984. 90 с.
- Н. Ф. Талызина. Управление процессом усвоения знаний: (Психологические основы). — 2-ое изд., доп. и испр. — М.: изд-во Моск. ун-та, 1984. — 334 с.
- В. Г. Ткачук. Современные концепции управления движениями // Лекция для студентов ИФК и слушателей факультетов повышения увалификации, преподавателей физ. воспитания и тренеров. — Киев, КГИФК, 1986. — 18 с.
- О. П. Топышев. Педагогические аспекты совершенствования деятельности спортсменов в игровых видах спорта: Автореф. дисс. ... д-ра пед. наук. — М., 1989. — 49 с.
- В. С. Фарфель. Управление движениями в спорте. — М.: Физкультура и спорт, 1975. — 208 с.
- А. Г. Фельдман. Центральные и рефлекторные механизмы управления движениями. — М.: Наука, 1975. — 183 с.
- Физиология мышечной деятельности: Учебник для ин-тов физкультуры / Под ред. Я. М. Коца. М.: Физкультура и спорт, 1982. 444 с.
- Ю. П. Фролов. Мозг и Труд. — М.: Медгиз, 1960. — 183 с.
- Л. В. Чхаидзе. Об управлении движениями человека. — М.: Физкультура и спорт, 1970. — 136 с.
- А. И. Шамардин. Исследование игровой деятельности вратаря в футболе и экспериментальное обоснование методики подготовки: Автореф. дис. ... канд. пед. наук. — М., 1979. — 15 с.
- Ю. Т. Шапков. Афферентный контроль произвольных движений // Тез. докл. всесоюз. науч.-практ. конф. / Физиологические механизмы целенаправленной деятельности спортсменов (Краснодар, 15-19 сентября, 1991). — М., 1991. — С. 80 — 82.
- Шейх Мульби Бен Саад. Влияние времени прослеживания мяча на двигательную структуру: Автореф. дисс. ... канд. пед. наук. — М., 1987. — 22 с.
- М. П. Шестаков, В. М. Зубков. «Аксон» — интеллектуальная компьютерная система планирования физической подготовки легкоатлетов — ТиПФК, 1994. — 8. — С. 35.
- A. E. Barts. Eye Movement latency duration and Response Time as a Function of Angulas Displacement // Journal of Experimental Psychology. — 1962. — V. 64. — P. 318 — 324.
- Beaugonin, M. and Haug, E., 1996, A numerical model of the human ankle/foot under impact loading in inversion and eversion. Society of Automotive Engineers, 962428, 239 — 249.
- P. M. Fitts. Perceptual — motor skills learning. In: Categories of human learning. N. Y. — London: 244, 1964. — 198 р.
- P. M. Fitts, B. K. Radford. Information capasity of dickrete motor responses under different cogninive sets. Journal of Experimental Psychology, 71, 4: 475. 1966. — Р. 98 — 114.
- P. Komi. Neuromuscular perfomance: Factors influencing force and speed production. — Scand. J. Sports Sci., 1979, 1, p. 2 — 15.
- P. V. Komi. Fatigue and recovery of neuromuscular function. Med. Sports Sci., 1984, 17, p. 187 — 201.
- S. Plagenhoef, 1972, Patterns of Human Motion — a Cinematographic Analysis, (N.J.: Prentice Hall), p. 89 — 116.
- E. C. Poulton. Measuring the Order of Difficulty of Visual Motor Tasks // Ergonomics. — 1958. — V. 1. — P. 234 — 239.
- M. Ridenour. Influence of ball sire speed direction height and distanse on interceptions of moving object // The Res. Quart. — 1977. — V. 48, № 1. — P. 138 — 143.
- R. H. Sharp. Keep the Eye on the Ball // Paper Presented at the Meeting of the British Society of Sport Psychology. — Leeds, 1972. — P. 32 — 37.
- Watts, R. G. and Ferrer, R., 1987, The lateral force on a spinning sphere. American Journal of Physics, 55, pp. 40-45.